PRINCIPIOS DEL ENTRENAMIENTO DE LA FUERZA.
Sobrecarga o de incremento progresivo de la carga: Es necesario aumentar el estímulo de una carga en forma progresiva.
Versatilidad: La variedad del entrenamiento es necesaria para mejorar las respuestas al entrenamiento y que no decaigan la motivación y las ansias de superación.
De individualidad: Cada deportista responde de forma diferente a cada carga.
De especificidad: El entrenamiento de la fuerza debe ser elegido convenientemente con las demandas de cada deporte, teniendo en cuenta.
1- El sistema energético dominante en cada deporte.
2- Los grupos de músculos específicos involucrados.
“Las capacidades de fuerza que se manifiestan directamente en los esfuerzos físicos (motrices) de magnitud dependen de la reacción integral del organismo que incluye la movilización de las cualidades y funciones psíquicas de todos sus sistemas fisiológicos. Por ello no hay que reducir las manifestaciones de fuerza del deportista a un concepto utilitario de fuerza de los músculos, es decir, sólo a las características mecánicas de sus propiedades contráctiles. Además hay que tener en cuenta, en primer lugar, que la fuerza muscular, al ser un componente dinámico de cualquier movimiento deportivo, puede tener diversas características cualitativas en función de su velocidad, la resistencia externa y la duración del trabajo. En segundo lugar, en condiciones de actividad deportiva el efecto de trabajo de los movimientos viene determinado tanto por la magnitud del esfuerzo máximo desarrollado como por el tiempo consumido para ello. El incremento de la fuerza muscular está determinado principalmente por el desarrollo de los cambios en la adaptación en el nivel del sistema nervioso central que llevan a una intensificación de la capacidad de los centros motores de movilizar un gran número de motoneuronas y de perfeccionar la coordinación intermuscular. El proceso de la contracción muscular está caracterizado por la secuencia de activación de las unidades motoras.
La fisiología investiga y discute sobre la correlación, en diferentes niveles, de dos mecanismos de regulación de la fuerza de la contracción muscular: la variación de las motoneuronas reclutadas y la frecuencia de su impulso. Algunos autores llegan a la conclusión de que el crecimiento de la fuerza en la fase inicial de la contracción se realiza, sobre todo, gracias al reclutamiento y después, a medida que crece la fuerza, el papel de reclutamiento disminuye y toma el papel principal el aumento de la frecuencia de impulso.
Desde el punto de vista psicológico en especial es necesario referirse a los esfuerzos volitivos, el estado psíquico del deportista y la concentración como elementos decisivos en la realización óptima de los ejercicios de fuerza. Es la perfección de la coordinación neuromuscular lo que posibilita la utilización de todo el potencial durante los máximos esfuerzos volitivos.
Con relación a este último factor, es importante referirse a que los más reconocidos científicos y entrenadores, señalan que aún cuando el atleta se encuentra en su mejor forma de preparación física y técnico-táctica lo manifestará en la práctica, en las competiciones en no más allá del 85 % de sus posibilidades aproximadamente y ello aumenta a medida en que la preparación psicológica del atleta sea más perfecta, logrando entonces el límite de sus posibilidades.
Un elemento que por lo general atletas y entrenadores no valoran y que es de gran importancia dentro de la ejecución de la fuerza es el llamado momento respiratorio.
La respiración, al realizar el ejercicio de fuerza, requiere de una regulación especial. Es sabido que los esfuerzos límites son factibles solamente con el pujo que es la tensión de la musculatura de la expiración al tener cerrada la glotis. El pujo eleva los índices de fuerza aunque solo debe acudirse a ello cuando sea necesario, es decir para soportar tensiones máximas muy buenas.
1.4 Factores bioquímicos y fisiológicos de los que depende la fuerza muscular.
“Toda actividad física exige energía biológica y su consumo está en correspondencia con el esfuerzo que se realice, la fuente de energía biológica durante la actividad muscular es la disociación del ATP, cuyo contenido en los músculos que trabajan es siempre relativamente constante, su recuperación ocurre de manera interrumpida (resíntesis) durante los procesos de reacciones químicas aeróbicas y anaeróbicas.
En los entrenamientos de fuerza las proteínas musculares son sometidas a grandes cambios, lo que implica durante el período de recuperación la supercompensación de estos, la cual se traduce en aumento de la masa muscular y una mayor disponibilidad de ATP.
El aporte energético de los esfuerzos de corta duración y de gran potencia se produce fundamentalmente por la vía del proceso anaeróbico aláctico. En este caso, la resíntesis del ATP dividido como resultado de la actividad muscular tan solo puede ser aportada si se emplean las reservas internas de fosfocreatina. Sin embargo, las exigencias de aporte energético del trabajo de fuerza no se limitan tan solo a esta fuente. El aumento de la potencia del sistema de glucolítico y glucólisis es característico de la adaptación a las grandes cargas de fuerza y corta duración. En los esfuerzos de fuerza máxima no superior a 6 segundos el lactato no se manifiesta ni en los músculos ni en la sangre, pero en los de 30 segundos su concentración aumenta de forma significativa, lo cual da testimonio de la incorporación del mecanismo glucolítico de aporte de energía.
Desde el punto de vista fisiológico la magnitud de la manifestación de la fuerza muscular durante la realización de ejercicios físicos depende, en primer término, de la formación de los reflejos condicionados que garanticen una gran concentración de los procesos de inhibición y excitación, además una contracción óptima y coordinada de los músculos necesarios en el funcionamiento de la resistencia dada.
En la contracción del músculo no participan todas las fibras musculares que lo componen por lo cual mientras mayor sea la excitación, mayor será la cantidad de fibras que participaron (Bowers y Fox, 1995).
El aumento del volumen muscular se efectúa por el espesamiento del diámetro y de la longitud de las miofibrillas. Con el entrenamiento de fuerza aumentan su diámetro sobre todo las fibras FT más allá del 25 % de la fuerza isométrica máxima de las fibras hay una solicitud selectiva Karlsson, (1975) Las fibras FT contribuyen al valor de la fuerza muscular en proporción mucho mayor que las fibras ST (Gollnick 1972, Buhrle y Schmitblekher, 1977).
1.5 Diferencia entre las fibras musculares, efectos del entrenamiento.
En investigaciones se han podido constatar diferencias fundamentales desde el punto de vista molecular entre ambos tipos de fibra, incluso dentro del mismo grupo, la subdivisión surge, no sólo como producto de preincubaciones químicas a distintos niveles de pH, sino también por la distinta estructura molecular.
Las diferencias en este último aspecto, se aprecian específicamente a nivel del sarcómero, particularmente en los filamentos gruesos de miosina. No se han apreciado deferencias entre las fibras I y II con relación a los filamentos de actina Billeter, (1981).
Teniendo en cuenta los distintos niveles de rendimiento en el deporte, dicha diferenciación surge, entre otros factores, debido a los tipos de encadenamientos de las cadenas peptídicas de la miosina. Las mismas se dividen entre cadenas pesadas y livianas. Las primeras son de tres tipos:
Cadena Pesada Rápida A (HCA, Heavy Chain A)
Cadena Pesada Rápida B (HCB, Heavy Chain B)
Cadena Pesada Lenta (HCS, Heavy Chain Slow)
Por su parte, las cadenas livianas de la miosina se identifican mediante la siguiente clasificación:
Cadena Liviana Rápida 1 (LCF1, Light Chain Fast 1)
Cadena Liviana Rápida 2 (LCF2, Light Chain Fast 2)
Cadena Liviana Rápida 3 (LCF3, Light Chain Fast 3)
Cadena Liviana Lenta 1 (LCS1, Light Chain Slow 1)
Cadena Liviana Lenta 2 (LCS2, Light Chain Slow 2)
Existen diferencias en los pesos moleculares entre las cadenas pesadas y las livianas; las primeras tienen un peso molecular de 200000, mientras las livianas varían entre 14600 y 23900. Las cadenas livianas se encuentran ubicadas en la cabeza de la miosina, lo cual es de gran importancia debido a que, precisamente en este lugar se encuentra ubicada la ATPasa, responsable en determinar el deslizamiento de la miosina con respecto a la actina. Las fibras musculares IIa y IIb tienen las mismas tres cadenas livianas rápidas (LCF1, LCF2, LCF3); esto induce a pensar que la sutil diferencia entre ambas puede estar en las dos cadenas pesadas, que obviamente son rápidas. Concretamente las diferencias se dan a través de distintas secuencias de aminoácidos primarios, las cuales se evidencian en diferentes niveles de la ATPasa durante su coloración histoquímica.
Las diferencias estructurales entre las fibras I con respecto a las II son muy evidentes, no sólo desde el punto de vista cualitativo sino también cuantitativo. Las fibras de contracción rápida II no poseen cadenas pesadas lentas, sólo rápidas, lo mismo ocurre en relación con las cadenas livianas. La cadena pesada de la fibra I, en cambio, es lenta; las cadenas livianas poseen no solamente cadenas livianas lentas, sino también rápidas su número varía y ello determinaría su distinta funcionabilidad Billeter, (1981). Se han podido encontrar variadas combinaciones de cadenas livianas lentas y rápidas; en algunos casos se encontraron solamente dos cadenas livianas lentas y dos rápidas. Hay que destacar, nuevamente, que esta variada estructura molecular determina distintos tipos de fibras I, Schanz, (1986).
En las actuales divisiones de fibras musculares también se hace mención a las IIc. ¿Cuáles son sus características? Las mismas no son únicamente de contracción rápida, sino además son resistentes debido a que poseen elevada densidad mitocondrial (Howald, 1989). Desde un punto de vista molecular, se encontró en estas fibras una amplia combinación de características propias de las fibras IIa, IIB y I. En las fibras IIc, la miosina posee dos cadenas pesadas, una lenta y otra rápida: HCF + HCS. Además de ello, existen tres cadenas livianas rápidas y dos cadenas livianas lentas. Estas características estructurales hacen de las fibras IIc las más versátiles desde el punto de vista funcional, en relación con las restantes. Sin embargo, entre las distintas biopsias efectuadas solo se han encontrado en una proporción del 2% con respecto a las restantes Billeter, (1981). La explicación de este fenómeno, es que las fibras IIc podrían constituir el paso intermedio entre las fibras I y II (en ambas direcciones y como producto de un proceso de entrenamiento).
1.6 Cambios entre los distintos tipos de fibras musculares.
Hay que reconocer en este aspecto que la fibra muscular posee un óptimo grado de plasticidad superior a los que en décadas pasadas sostuvo Nett, (1955). Es indudable que, para entonces, se consideraron únicamente cambios funcionales sumamente globales, los que se podrían apreciar únicamente por un mejor rendimiento deportivo. Para que se constaten reales variaciones en estos cambios, es necesario realizar extensos estudios longitudinales que abarquen desde meses hasta, quizás, años de entrenamiento muy específicos. De todos modos, con un entrenamiento sistemático que abarque algunas semanas, se constatan íntimos cambios estructurales como también metabólicos.
Estos cambios se producen desde el tipo de fibras II al I, pero no a la inversa Jolesz, Sreter, (1981), sin que estos cambios enzimáticos o moleculares impliquen una transformación histoquímica del tipo de fibra I o II. Las transformaciones se pueden resumir de la siguiente forma (teniendo en cuenta la aplicación de estímulos lentos):
· La actividad enzimática oxidativa y la densidad capilar pueden incrementarse con una semana de trabajo;
· Con dos semanas de entrenamiento se producen cambios en la propiedad contráctil isométrica;
· En la tercera un descenso de la actividad de la ATPasa.
· Entre la tercera y sexta semana existen modificaciones a escala molecular con respecto a la estructura de la miosina y la troponina (Salmons, Henriksson, 1982).
Esto en lo que se refiere a esfuerzos desarrollados en forma relativamente “lenta”, a la manera de la metodología aeróbica, pero no corriendo, sino aplicando estímulos eléctricos de baja frecuencia (10 impulsos / seg.) y durante un período relativamente prologado. ¿Ocurre lo mismo en el camino inverso? ¿Pueden las fibras I adoptar las características del grupo II? Los hechos y comprobaciones científicas están demostrando que no. Aparentemente, en un primer momento, existe una transformación; las fibras I se hacen “más veloces”, pero no bien cesa el estímulo, estas retornan a sus características genéticas originales (Salmons, Henriksson, 1982). Esto lo comprendemos desde el momento en que las actividades de la vida diaria de relación constituyen, básicamente, estímulos de bajo umbral de excitación, las cuales están en consonancia con las fibras de contracción lenta: las fibras I están diariamente en su verdadero medio ambiente. Observaciones como estas nos inducen a pensar que los deportistas especialistas en velocidad podrían mejorar sensiblemente su potencia y capacidad aeróbica mediante entrenamientos sistemáticos efectuados a la manera de un fondista, por el contrario, el camino inverso parece improbable.
1.7 Cambios metabólicos causados por el entrenamiento.
Las modificaciones que pueden tener algunos tipos de fibras, son producto de aquellos cambios que sufren como consecuencia de sistemáticos estímulos específicos a los cuales se someten. Es por este motivo que, estudios histoquímicos y electro microscópicos han podido constatar interesantes transformaciones en las fibras como producto del entrenamiento sistemático. Además, es interesante que una misma clase de estímulo puede producir diferentes resultados, según el grupo de fibra muscular de que se trate; en otros casos, en cambio, ambos tipos de fibras tienen cambios similares, pero con estímulos de distintas características. El volumen mitocondrial se incrementa en gran magnitud como producto de entrenamientos sistemáticos, a la manera de los fondistas. Holloszy (1971), encontró un incremento del 70% del contenido proteico mitocondrial por gramo de músculo, como también un incremento del 100% de la capacidad oxidativa, en caballos entrenados con esfuerzos prolongados. El incremento del volumen mitocondrial se da tanto en las fibras I como en las II; sin embargo los más llamativos de ello son estos cambios que se dan en mayor magnitud en las fibras II con respecto a la I. Además se produce una elevada adaptación a escala enzimática, con incremento de su actividad. Esfuerzos aeróbicos de baja intensidad modifican la actividad enzimática de las fibras I y el mismo resultado se comprueba en las fibras musculares rápidas II, pero en este último caso, mediante la combinación de trabajos anaeróbicos (Henriksson, Reitmann, 1976). Las enzimas que incrementan su actividad son las oxidativas, tales como succinato deshidrogenasa (SDH), succinato oxidasa (SO) y la citocromo oxidasa (Holloszy, 1971; Howald, 1989). Esto podría apoyar la hipótesis de que la resistencia y/o potencia aeróbica, no solamente se puede incrementar mediante prolongados esfuerzos de duración, sino también mediante el entrenamiento fraccionado rápido (aeróbico-anaeróbico). Usando esta metodología, se tienen muy especialmente en cuenta las fibras musculares II, de contracción rápida y sin desmedro de la velocidad como en el caso anterior. Por otra parte, los trabajos de fuerza muscular activan las enzimas citoplasmáticas tales como la mioquinasa, CPK, ATPasa y PFK. Las fibras musculares de contracción rápida son considerablemente más susceptibles a la hipertrofia que las STI, con mayor síntesis proteica a nivel ribosomal.
De acuerdo con ello cambia la relación FTF/STF con relación a su superficie y en beneficio de las fibras rápidas (FTF). Esto se ha comprobado en levantadores de pesas como también en lanzadores y saltadores (Edstrom y Ekblom, 1972; Pince, 1976).
En conclusión, la fibra muscular posee un elevado grado de adaptación, constituyéndose en excelente receptor para métodos específicos de entrenamiento. Sin embargo, también puede ocurrir que si no se entrena controlando efectivamente todos los indicadores de la carga, se modificará el metabolismo de la célula muscular y se altera su estructura, logrando una adaptación de fuerza en un sentido no deseado para el deporte y resultados deportivos inferiores a los que pudieron ser y por esta razón no se cumplan los objetivos de la especialidad deportiva solamente.
Esto lleva a concluir que la preponderancia metodológica del entrenamiento debería estar estructurada preferentemente, sobre el mecanismo energético y el régimen de trabajo muscular que caracteriza a la especialidad deportiva
Después de valorar los principios y algunas influencias fisiológicas, bioquímicas y psicológicas relacionadas con el entrenamiento de la fuerza, es importante tener en cuenta antes de realizar un análisis de los métodos de entrenamiento, algunos elementos relacionados con la fuerza y la velocidad del movimiento cuando se trabaja con una carga o resistencia externa.
1.8 FUERZA TIEMPO Y FUERZA VELOCIDAD.
A nivel fisiológico las tensiones producidas por el sistema neuromuscular son la fuente de todo movimiento voluntario y determinan el nivel de fuerza aplicada, la velocidad y potencia aplicada expresada durante un trabajo, pero desde el punto de vista biomecánico es el análisis de la relación entre la producción de fuerza y el tiempo lo que expresa adecuadamente las formas en que el sistema neuromuscular trasmite fuerza.
El adecuado análisis de la producción de fuerza en relación con tiempo es esencial para determinar y controlar la evolución de las características funcionales de los sujetos al aplicar las distintas formas en que se manifiesta la fuerza muscular. Ya que las diversas formas que adquiera la curva fuerza-tiempo reflejan el patrón de activación neuromuscular por el cual cada sujeto produce la fuerza necesaria para realizar un movimiento a una velocidad potencia determinada.
El nivel porcentual de fuerza producida en un tiempo dado son los datos de referencia más importantes en torno a los cuales se estiman las características funcionales de cada sujeto. Badillo (2003), Nalclerio (2001).
Indicadores relacionados con estas curvas:
· Impulso de la fuerza que se expresa por toda el área que se encuentra bajo la curva.
· Valor de la fuerza máxima y el tiempo en que se tarda en producirla.
· Magnitud de fuerza inicial que refleja la habilidad del sistema neuromuscular para generar altos niveles de fuerza en los primeros instantes de activación (partiendo desde un nivel de tensión = 0), lo cual se refleja en la curva en los primeros 100 milisegundos (Mls). En la práctica, índice de fuerza inicial se estima a partir del coeficiente entre la magnitud de fuerza producida a nivel del 50 % de la fuerza máxima isométrica y el tiempo.
· Tiempo necesario para alcanzar un nivel determinado de fuerza isométrica o dinámica. Este valor expresa el índice de fuerza explosiva el cual puede medirse de diferentes maneras.
Hasta generar un porcentaje de fuerza respecto a la máxima posible, por ejemplo el tiempo en que se tarde en generar el 50 % de la fuerza máxima.
Hasta generar un valor determinado de fuerza en términos absolutos.
Estos índices se pueden tomar tanto en el inicio de la curva como en cualquier parte de esta, según sean las necesidades del investigador.
En la mayoría de los gestos deportivos, especialmente aplicados al trabajo con pesas, la relación entre la generación de fuerza y el tiempo invertido en producirla, es en gran medida determinado por la magnitud de la carga a vencer, ya que existe una relación positiva casi perfecta entre la carga desplazada y la cantidad de fuerza en newton que se puede generar, lo cual es especialmente apreciado al analizar la producción de fuerza aplicada en ejercicios con pesas al movilizar cargas de diferentes magnitudes a la mayor velocidad posible.
El espectro que expresa la relación entre el nivel de fuerza aplicada con diferentes magnitudes de carga y el tiempo necesario para producirlo, muestra cómo el nivel de carga a desplazar determina las características funcionales del sistema neuromuscular, ya que la capacidad de aplicar altos niveles de fuerza cae a medida que las cargas van disminuyendo, aunque el pico máximo se alcance antes.
Así, cuando más baja es la carga, menos posibilidades se tendrán de producir altos niveles de fuerza, pero esta podrá desarrollarse más rápidamente.
El análisis de la curva fuerza tiempo (F-T) permite comparar la evolución de las manifestaciones
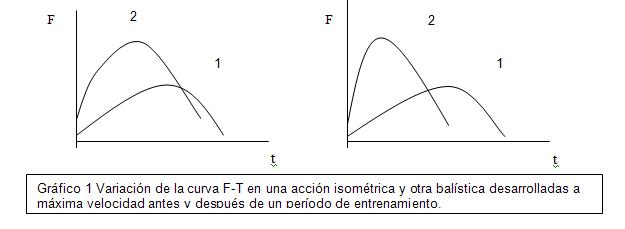
de fuerza a lo largo de un período de entrenamiento o comparar las características funcionales entre diferentes sujetos, así por ejemplo en la gráfica 1 se muestra cómo se puede modificar la curva producida antes (1) y después (2) de un período de fuerza explosiva, donde el mismo sujeto alcanza niveles más altos en menos tiempo, tanto en una acción isométrica (izquierda), donde se aplica fuerza máxima a la velocidad con un gesto balístico, (derecha) como por ejemplo un lanzamiento.
Al comparar a dos sujetos que expresan el mismo nivel de fuerza máxima y analizar la forma en que cada uno se expresa en diferentes niveles porcentuales, podría suceder que uno llegara más rápido que el otro a un nivel de fuerza dado, por ejemplo el 30 % por lo cual este tendrá ventajas sobre el otro en actividades donde se requiera una producción de fuerza en corto espacio de tiempo, por ejemplo en los saltos, pero puede que estas ventajas no sean importantes si los dos atletas midieran solamente la capacidad para generar fuerza máxima absoluta a través de un clásico test de carga máxima con pesos (test de 1MR). Si dos sujetos tuvieran la capacidad de generar altos niveles de fuerza a un porcentaje dado, por ejemplo al 50 % de la fuerza máxima isométrica, pero uno tuviera más fuerza máxima, este tendría ventajas ya que el valor absoluto alcanzado al 50 % se corresponderá con un nivel absoluto en Kg. o Newton mayor.
El análisis de la curva fuerza tiempo refleja no solo las características funcionales del sujeto sino cómo este se adapta a las cargas de entrenamiento, lo cual es analizado al relacionar la producción de fuerza con la velocidad generada, ya que esta refleja el tiempo en que se tarda en recorrer en determinado espacio o rango de movimiento
Velocidad = espacio recorrido/ tiempo invertido en recorrerlo
Análisis de la relación fuerza velocidad.
La relación existente entre el valor de fuerza aplicado y la velocidad generada (R F-V) ha sido analizada originalmente por la H.V. Hill (citado por Martín, 2001) en sus estudios de laboratorio con músculos aislados, determinando una relación inversa hiperbólica entre la F-V, donde se observa una hipérbola con acintoticas paralelas a los dos ejes principales (X;Y), es decir que ambos extremos de la línea, la relación fuerza velocidad se va haciendo paralelo al eje, de modo que solo conserva una relación lineal en la zona central.
Esta relación entre la magnitud de fuerza aplicada y la velocidad de movimiento observada, en diferentes regímenes de trabajo muscular (isocinético, dinámico concéntrico, excéntrico o gestos con ciclos de estiramiento acortamiento) difiere notablemente de la reportada inicialmente por Hill e incluso es diferente entre los distintos regímenes de trabajo muscular que pueden manifestarse en los trabajos de fuerza, lo que se hace más evidente al comparar la relación F-V manifestada respecto a los niveles porcentuales de carga máxima en movimientos dinámicos concéntricos con cargas naturales o régimen isoinercial que es la forma más habitual de realizar. Los ejercicios de fuerza presentan una relación negativa casi perfecta entre el nivel de fuerza y velocidad de movimiento, lo cual difiere de la reportada en el régimen isocinético en los que se denota una forma hiperbólica similar a la reportada por Hill, pero con una relación más parabólica en los extremos.
Al analizar la relación fuerza velocidad y considerar que la potencia expresada durante un ejercicio resultado del producto entre la fuerza aplicada y la velocidad generada, al relacionar estas dos variables (fuerza y velocidad) también pueden determinarse los niveles resultantes de potencia en cada caso.
La relación entre la fuerza, velocidad y potencia reportada en los trabajos originales de Hill en músculos aislados mostró que la máxima expresión de potencia se alcanza con el 30% de la máxima velocidad de acortamiento muscular y con un 25 % de la fuerza máxima isométrica, pero como hemos visto en los ejemplos anteriores, el patrón de comportamiento mecánico de un músculo aislado no se puede transferir hacia el patrón a realizar en movimientos musculares en los seres vivos (Hombre) ya que en estos la relación entre fuerza-velocidad-potencia se expresa de forma diversa, dependiendo no solo del régimen de trabajo muscular, sino de otros factores como son, la mecánica específica del ejercicio valorado y factores que atañen a la individualidad de cada sujeto, como nivel de entrenamiento, su configuración antropométrica y porcentaje de fibras rápidas.
La valoración reiterada de la curva de fuerza-velocidad-potencia a lo largo de un período de entrenamiento permite no solo conocer las tendencias fisiológicas del sujeto sino también medir los cambios producidos sobre el comportamiento motor del deportista, lo que constituye un elemento de diagnóstico, control y planificación del entrenamiento, ya que ofrece información sobre los verdaderos cambios funcionales y estructurales que se van determinando en el organismo, citado por Naclerio 2001.